La concentración de adenosin trifosfato, la "moneda de cambio" molecular de energía, debe ser muchas veces superior a la de ADP (di-fosfato). Si usas la energía de un enlace de fósforo, la molécula queda solo con dos. Así aumenta la concentración de ADP, lo que a la larga limita el consumo de ATP hasta detenerlo. Y el ciclo de formación de ATP comienza de nuevo....
¿paralelismos con los mercados financieros, suma de decenas de inversiones fallidas para conseguir inversiones rentables?
Pep Sandoval
"Las células mantienen la proporción de ATP a ADP en el punto de diez órdenes de magnitud del equilibrio, siendo las concentraciones de ATP miles de veces superior a la concentración de ADP. Este desplazamiento del equilibrio significa que la hidrólisis de ATP en la célula libera una gran cantidad de energía. Al ATP se le llama a veces "molécula de alta energía", aunque esto no es correcto, ya que una mezcla de ATP y ADP en equilibrio en el agua no puede hacer un trabajo útil".
http://www.coenzima.com/adenosina_trifosfato_atp
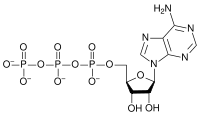
Adenosina trifosfato (ATP)
 |
El ATP fue descubierto en 1929 por Karl Lohmann. En 1941, Fritz Albert Lipmann propuso el ATP como principal molécula de transferencia de energía en la célula.
PROPIEDADES Y ESTRUCTURA
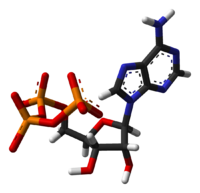 |
El ATP es altamente soluble en agua y muy estable en soluciones de pH entre 6.8 y 7.4, pero se hidroliza rápidamente a pH extremo. Por consiguiente, se almacena mejor como una sal anhidra.
La masa molecular del ATP es de 507,181 g/mol y su acidez es de 6.5. Es una molécula inestable y tiende a ser hidrolizada en el agua. Si el ATP y el ADP se encuentran en equilibrio químico, casi todos los ATP se convertirán a ADP. Las células mantienen la proporción de ATP a ADP en el punto de diez órdenes de magnitud del equilibrio, siendo las concentraciones de ATP miles de veces superior a la concentración de ADP. Este desplazamiento del equilibrio significa que la hidrólisis de ATP en la célula libera una gran cantidad de energía. Al ATP se le llama a veces "molécula de alta energía", aunque esto no es correcto, ya que una mezcla de ATP y ADP en equilibrio en el agua no puede hacer un trabajo útil. El ATP no contiene "enlaces de alta energía", y cualquier otra molécula inestable serviría como una forma de almacenar energía si la célula mantuviera su concentración lejos del equilibrio.
El ATP tiene múltiples grupos ionizables con diferentes constantes de disociación del ácido. En solución neutra, el ATP está ionizado y existe principalmente como ATP4-, con una pequeña proporción de ATP3-. Como tiene varios grupos cargados negativamente en solución neutra, puede quelar metales con una afinidad muy elevada. El ATP existe en la mayoría de las células en un complejo con Mg2+.
FUNCIONES
Fuente de energía
El ATP es la principal fuente de energía para la mayoría de las funciones celulares. Esto incluye la síntesis de macromoléculas como el ADN, el ARN y las proteínas. También desempeña un papel fundamental en el transporte de macromoléculas a través de las membranas celulares, es decir, en la exocitosis y endocitosis.
Debido a la presencia de enlaces ricos en energía (entre los grupos fosfato son los enlaces anhídrido del ácido), esta molécula se utiliza en los seres vivos para proporcionar la energía que se consume en las reacciones químicas. De hecho, la reacción de hidrólisis de la adenosina trifosfato en adenosina difosfato y fosfato es una reacción exergónica donde la variación de entalpía libre estándar es igual a -30,5 kJ/mol:
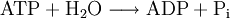
Por el contrario, la reacción de síntesis de la adenosina trifosfato a partir de adenosina difosfato y fosfato es una reacción endergónica donde la variación de entalpía libre estándar es igual a +30,5 kJ/mol:

La reacción de hidrólisis del ATP en adenosín monofosfato (y pirofosfato) es una reacción exergónica donde la variación de entalpía libre estándar es igual a -42 kJ/mol:
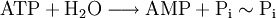
La energía se almacena en los enlaces entre los grupos fosfato.
Sin embargo, hay un nivel de entalpía a sobrepasar antes de liberar esta energía (estado de transición). Esto explica por qué la hidrólisis de los enlaces pirofosfato no sucede todo el tiempo. Las enzimas son capaces de reducir ese umbral de entalpía para utilizar la energía liberada.
Si la energía se almacena en los enlaces anhídridos, podríamos preguntarnos cuál es el interés de los seres vivos para sintetizar la molécula en su conjunto y no sólo el pirofosfato libre. La razón es, probablemente, la capacidad de las enzimas para reconocer el ATP, más fácil de hidrolizar específicamente que los pirofosfatos libres, que son muy similares a todos los grupos fosfatos presentes en las biomoléculas.
El ADP puede ser fosforilado por la cadena respiratoria de las mitocondrias y los procariotas, o por los cloroplastos de las plantas, para restaurar el ATP. La coenzima ATP/ADP es un proveedor de energía universal, y es la principal fuente de energía directamente utilizable por la célula. En los seres humanos, el ATP constituye la única energía utilizable por el músculo.
En la síntesis del ácido nucleico ARN, el ATP es uno de los cuatro nucleótidos incorporados directamente en las moléculas por las enzimas ARN polimerasas. La energía que conduce esta polimerización procede de la ruptura del pirofosfato (dos grupos de fosfato). El proceso es similar en la biosíntesis de ADN, salvo que el ATP se reduce al desoxirribonucleótido dATP, antes de su incorporación en el ADN.
El ATP está críticamente involucrado en el mantenimiento de la estructura celular, facilitando el montaje y desmontaje de elementos del citoesqueleto. En un proceso similar, el ATP es necesario para el acortamiento de los filamentos de actina y miosina necesarios para la contracción muscular. Este último proceso es una de las principales necesidades energéticas de los animales y es esencial para la locomoción y la respiración.
Señalización extracelular
El ATP, el ADP o la adenosina son reconocidos por los receptores purinérgicos. En los seres humanos, esta señalización tiene un importante papel tanto en el sistema nervioso central como en el periférico. La liberación de ATP de las sinapsis, los axones y la neuroglía activa los receptores de membrana purinéricos conocidos como P2. Los receptores P2Y son metabotrópicos, es decir, modulan el calcio intracelular y, a veces, los niveles de AMP cíclico.
Señalización intracelular
Es utilizado por las quinasas como la fuente de grupos fosfato en sus reacciones de transferencia de fosfato. La actividad de las quinasas sobre los sustratos como las proteínas o los lípidos de la membrana son una forma común de transducción de señales. La fosforilación de una proteína por una quinasa puede activar esta cascada.
La adenilato ciclasa también usa el ATP y lo transforma en AMP cíclico (AMPc), una molécula segundo mensajero que está involucrada en el desencadenamiento de las señales de calcio mediante la liberación de calcio intracelular. Esta forma de transducción de señales es particularmente importante en la función cerebral, aunque está involucrada en la regulación de multitud de otros procesos celulares.
Síntesis de desoxirribonucleótidos
En todos los organismos conocidos, los desoxirribonucleótidos que componen el ADN se sintetizan por la acción de enzimas ribonucleótido reductasas (RNR). Estas enzimas reducen el grupo hidroxilo 2' en el azúcar ribosa, que pasa a ser desoxirribosa, formando un desoxirribonucleótido (dATP). Todas las enzimas ribonucleótido reductasas usan un radical sulfidrilo común en un mecanismo de reacción que depende de los residuos cisteína, que se oxidan para formar enlaces disulfuro en el curso de la reacción. Las enzimas RNR son recicladas mediante reacción con tiorredoxina o glutaredoxina.
ALMACENAMIENTO DE ATP
Las reservas de ATP en el organismo no exceden de unos pocos segundos de consumo. En principio, el ATP se produce de forma continua, pero cualquier proceso que bloquee su producción provoca la muerte rápida (como es el caso de determinados gases de combate diseñados para tal fin; o venenos como el cianuro, que bloquean la cadena respiratoria; o el arsénico, que sustituye el fósforo y hace que sean inutilizables las moléculas fosfóricas).
Las moléculas de creatina enlazan un fosfato mediante un enlace rico en energía como el ATP. El ADP puede convertirse en ATP por acoplamiento con la hidrólisis de fosfato de creatina. La creatina, por tanto, recicla el fosfato liberado por la hidrólisis de la molécula de ATP original. Esto ayuda a mantener la energía fácilmente movilizada sin agotar las reservas de ATP.
El ATP no se puede almacenar en su estado natural, sino sólo como intermediarios de la cadena de producción de ATP. Por ejemplo, el glucógeno puede ser convertido en glucosa y aportar combustible a la glucolisis si el organismo necesita más ATP. El equivalente vegetal del glucógeno es el almidón. La energía puede también ser almacenada como grasa, mediante neo-síntesis de ácidos grasos.
No hay comentarios:
Publicar un comentario